|
Our research interests are in neural development,
plasticity, and sensory processing. Nervous systems
require a high level of organization and precision, as
each neuron is itself a complex 3-dimensional entity that
may receive anywhere from 1 to over 300,000 inputs
(synapses) from other neurons. Individual neurons
interacting with other neurons to perform specific tasks
make up neural circuits. Within a neural circuit, the
type, location, and strength of each of the synapses
connecting the neurons determine the quality and range of
tasks the circuit can perform.
The precision achieved by neural
circuits is neither hard-wired nor randomly acquired.
Rather, this precision is attained through an interplay of
environmental and genetically-determined factors during an
early developmental period. While the specific task to be
performed and the timing of the early developmental period
vary with brain area, a common theme for circuit
refinement throughout the nervous system is that neural
activity (whether spontaneously generated in the
nervous system or driven by external stimuli) directs
biological mechanism (ie., specific proteins and
cellular processes) to modify neural circuits (plasticity). Neuroscientists
refer to this process as "activity-dependent plasticity."
Our aim is to understand the rules and mechanisms by which
immature neural circuits are refined, especially those
mechanisms related to activity-dependent plasticity and to
the coordinated refinement of excitatory and inhibitory
circuits.
We study how neural circuits are
refined during early life--and how circuit precision
contributes to sensory processing--in the mammalian
auditory system, with a current focus on the lateral
superior olive (LSO). This brainstem nucleus plays a
central role in auditory processing and is critical for
sound localization and extracting signal from noise, key
elements of auditory perception and attention. The LSO is
part of a larger auditory processing circuit, the superior
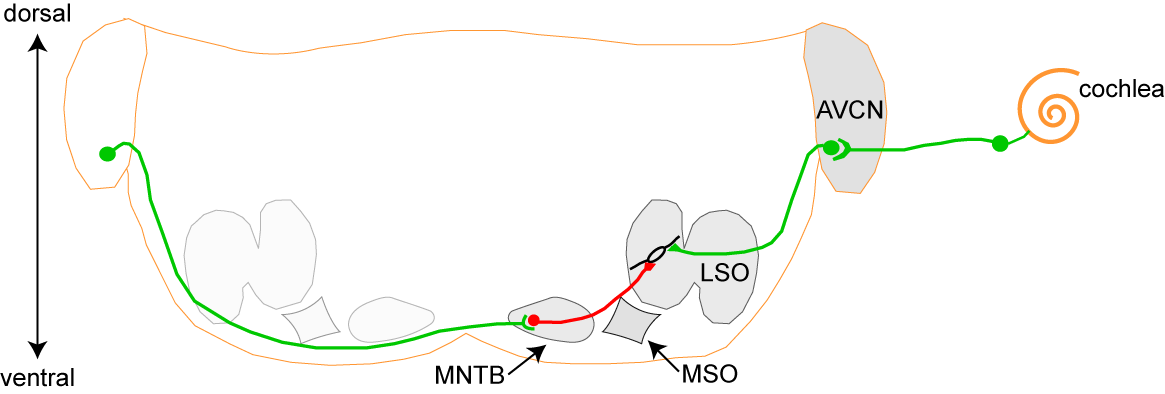
olivary complex (SOC), depicted below.
To localize sound sources along the
azimuth, many species use differences in sound arrival
time and intensity between the two ears; in mammals these
interaural differences in phase and intensity are computed
in the SOC. In particular, large neurons in the mammalian
LSO "add" two inputs that both encode sound
intensity: a positive (excitatory, glutamatergic)
input from the ear on the same side and a negative
(inhibitory, glycinergic) input from the other ear. To
reliably compute these interaural intensity differences,
the LSO cells require inputs with precise tonotopic
(frequency-based) alignment. A major task for the
developing brain is to establish and refine the "circuit
diagram" of the LSO so that individual LSO cells receive
inhibitory and excitatory inputs that correspond to the
same sound frequency.
We work at the interface of cellular
and systems neuroscience. Current research directions in
the lab concern the nature and function of GABA, glycine,
and glutamate co-transmission during development,
specification of synapse position during development,
information encoded in early spontaneous activity in the
auditory brainstem, and mechanisms of
developmental/synaptic plasticity in the LSO.
|
|
|
Like most neural structures, the auditory
brainstem is bilaterally symmetric; for clarity, only one
side is labeled here. The medial superior olive (MSO) is
primarily responsible for computing interaural timing
differences, the LSO for computing interaural intensity
differences. The major inputs to the LSO are the
excitatory (green) input from the ipsilateral
anteroventral cochlear nucleus (AVCN), a glutamatergic
projection, and the inhibitory (red) projection from the
ipsilateral medial nucleus of the trapezoid body (MNTB).
The inhibitory MNTB-LSO projection carries information
from the contralateral ear; though it is glycinergic in
the adult, it uses GABA, glycine, and glutamate during the
period of major circuit refinement.
|
|